 |
| Projects |
|
|
| Mutants
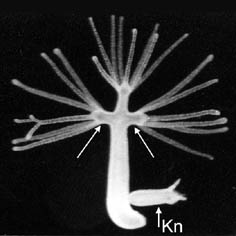
The mutant strain multi-headed-1 (mh-1) is a sexual inbreed generation of the wildtype (wt 105) of Hydra magnipapillata. It is able to produce several ectopic heads along its body axis (see figure, arrows).
By producing
chimeras of various cell types we could show that the cause for the
ability to form extra heads is located in the ectodermal epithelial
cells of mh-1. The interstitial cells are not involved in this
process. S. Zeretzke and S. Berking (1996) Analysis of a hydra mutant which produces extra heads along the body axis. Int. J. Dev. Biol. Suppl.1:271S S. Zeretzke and S. Berking (2001) Pattern regulation of a Hydra strain which produces additional heads along the body axis. Int. J. Dev. Biol. 45: 431-439 (2001) (PDF_File) S. Zeretzke and
S. Berking (2002) In the multiheaded strain (mh-1) of Hydra
magnipapillata the ectodermal cells are |
| home |
| Budding of polyps In Hydra sp. (Hydrozoa) and in Cassiopea sp. (Scyphozoa) a bud develops at the gastral region of the parent animal. The tissue of the bud does not originate from a meristem but from the gastral tissue of the parent animal. An important difference to note is that in hydrozoa the tip of the bud develops into the head of the polyp while in (many) scyphozoa the tip develops into the opposite part, namely the foot. Therefore, the central question is whether the systems which are controlling pattern formation in these animals are fundamentally different. A summary of the latest experimental results and of the disscusion about models of control mechanisms of bud development in hydra can be found under Berking, 1998. There is a recent paper addressing bud development in Cassiopea sp. We offer a theoretical model of pattern formation which allows to understand bud development in hydrozoa and scyphozoa. Publications : Berking, S.
(1977). Bud formation in Hydra: Inhibition by an endogenous morphogen.
Roux's Arch. Dev. Biol. 181, 215-225. Kehls, N.E., K.
Herrmann and S. Berking (1999) The protein phosphatase inhibitor
cantharidin induces head and foot formation in buds of Cassiopea
andromeda (Rhizostomae, Scyphozoa) Int. J. Dev. Biol. 43:51-58
|
| home |
| home |
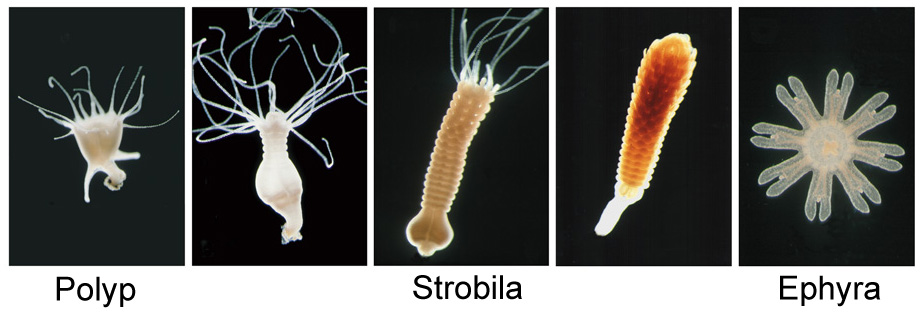
| Metamorphosis
and strobilation in Aurelia aurita
Abstract Metamorphosis Larvae of cnidarians need an external cue for metamorphosis to start. The larvae of various hydrozoa, in particular of Hydractinia echinata respond to Cs+, Li+, NH4+ and seawater in which the concentration of Mg2+-ions is reduced. They further respond to the phorbolester Tetradecanoyl-phorbol-13-acetat (TPA) and the diacylglycerol (DAG) diC8 which both are argued to stimulate a proteinkinase C. The only well studied scyphozoa, Cassiopea spp., respond differently, i.e. to TPA and diC8 only. We found that larvae of the scyphozoa Aurelia aurita, Chrysaora hysoscella and Cyanea lamarckii respond to all compounds mentioned. Trigonelline (N-methylnicotinic acid) a metamorphosis inhibitor found in Hydractinia larvae which is assumed to act by delivering a methyl group for transmethylation processes antagonises metamorphosis induction in Chrysaora hysoscella and Cyanea lamarckii. The three species tested are scyphozoa belonging to the subgroup of semaeostomeae, while Cassiopea spp. belong to the rhizostomeae. The results obtained may contribute to the discussion concerning the evolution of cnidarians and may help to clarify if the way metamorphosis can be induced in rhizostomeae as a whole is different from that in hydrozoa and those scyphozoa belonging to the subgroup semaeostomeae. Abstract Strobilation Polyps of Aurelia
aurita can transform into several medusae (jellyfish) in a process
of sequential subdivision. During this transformation, two processes
take place which are well known to play a key role in the formation of
various higher metazoa: segmentation and metamorphosis. In order to
compare these processes in bilaterians and cnidarians we studied the
control and the kinetics of these processes in Aurelia aurita.
Segmentation and metamorphosis visibly start at the polyp’s head and
proceed down the body column but do not reach the basal disc. The small
piece of polyp which remains will develop into a new polyp. The
commitment to the medusa stage moves down the body column and precedes
the visible onset of segmentation by about one day. Segmentation and
metamorphosis can start at the cut surface of transversely cut body
columns, leading to a mirror-image pattern of sequentially developing
medusae. Strobilation of Aurelia aurita
Publications: B Siefker, M Kroiher, S Berking (2000) Induction of metamorphosis from the larval to the polyp stage is similar in Hydrozoa and a subgroup of Scyphozoa (Cnidaria, Semaeostomeae). Helgoland Marine Research 54, pp 230-236 M Kroiher, B
Siefker, S Berking (2000) Induction of segmentation in polyps of Aurelia
aurita (Scyphozoa, Cnidaria) into medusae and formation of
mirror-image medusa anlagen. Int. J. Dev. Biol. 44, pp
485-490 Herrmann
K, Siefker B, Berking S (2003) Sterile poystyrene culture dishes induce
transformation of polyps into medusae in Aurelia aurita (Scyphozoa,
Cnidaria). Methods in Cell Science 25, 135-136 Berking S, Czech N,
Gerharz M, Herrmann K, Hoffmann U, Raifer H, Sekul G, Siefker B,
Sommerei A, Vedder F (2005) |
| home |
| home |
| Models
of control of pattern formation
To formulate theoretical models for control of pattern formation it is forst necessary to arrange a lot of experimental data with possible rare assumptions. We wish to understand the mechanisms of the control processes. We are aware that theoretical models are incomplete and preliminary. It is the function of such models to point the way for further experimental work and to help to establish the precise nature of the molecular and biochemical basis of control of pattern formation. From our point of
view pattern formation in cnidaria is hierarchically organized. The
primary system controls the positional value of the tissue, while the
secondary systems become active at distinct positional values and
determine the local differentiation, for example the formation of
tentacles or the formation of the basal plate. As we take it, there are
two types of dominance in the primary system. The type 1 dominance is
identical to that autocatalysis proposed by Gierer and Meinhardt (1972,
Kybernetik) and therewith the coupled inhibition of this autocatalysis
in the surrounding of autocatalytically active cells. In the area of
autocatalysis the positional value of the tissue increases, if not the
type 2 dominance is effective. The type 2 dominance is based on the
assumption that in the area of autocatalysis a further long range
signal
is generated, which decreases the positional value of the surrounding
tissue. Publications: Berking, S.
(1979b). Analysis of head and foot formation in Hydra by means of an
endogenous inhibitor. Roux's Arch. Dev. Biol. 186, 189-210.
|
| home |
| Separation
of buds
Hydra reproduces
preferently asexually through budding The buds were build in a distinct
region of the body colomn the so called budding region. The first
visible tip of the bud developes into the hypostom of the animal.
Then the gastric region of the bud grows untill the bud forms foot
tissue, builds a constriction and separates from the parent animal. We
can show that signaltransduction pathways are involved in the pattern
forming process at the bud´s base. Especially modulators of
protein kinases interfere with this process. (F. Pérez and S.
Berking, 1994, Roux´s Arch. Dev. Biol. 203: 284-289.).
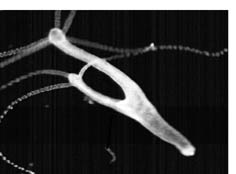
Abstract: The fresh water polyp Hydra can reproduce asexually by forming buds. These buds separate from the parent animal due to the developement of foot tissue in a belt-like region and the formation of a constriction basal to that region. A single pulse treatment with activators of protein kinase C , including 1,2-dioctanoyl-rac-glycerol and 1,2-o-tetradecanoylphorbol-13-acetate, and inhibitors of various protein kinases, including staurosporine, H-7 and genistein, interferred with foot and constriction formation. The buds did not separate. Therewith branched animals were formed some of which bore a lateral foot patch. Simultaneous treatments with an activator and a inhibitor led to a higher amount of branched animals than treatments with one of these agents alone. Based on the different specifities of the activators and the inhibitors used we propose that activation of a protein kinase C and/or inhibition of a probably non-C-type protein kinase interfere with the decrease of positional value at the bud´s base, a process necessary to initiate the pattern forming system leading to foot formation. Furthermore there exists a link that a serine / threonine protein phosphatase (Typ 2A) is involved in pattern formation at the bud´s base. Incubating the animals with cantharidin (inhibitor of the PP2A) leads to the inhibition of foot formation at the bud´s base. It is out of question that protein kinases and phosphatases, which have an important function as a part of signal cascades during developmental processes in various organisms, are also involved in pattern formation in Hydra, which is an very old organism. Besides serine /
threonine and tyrosin kinases and phosphatases we are interested in
ions
which are directly or indirectly connected with those kinases /
phosphatases and play a role in the foot formation at the bud´s
base. We focus especially calcium and lithium. We can show that a
dcrease of the calcium concentration in the culturemedium to the
intracellular level leads to the formation of branched animals (s.
figure). Publications: Hassel, M., and
Berking, S. (1990). Lithium ions interfere with pattern control in Hydra
vulgaris. Roux's Arch. Dev. Biol. 198, 382-388. Stefanie
Zeretzke, Fernando Pérez, Kirsten Velden and Stefan Berking
(2002) Ca2+-ions and pattern |
| home |
| home |
| home |
| home |
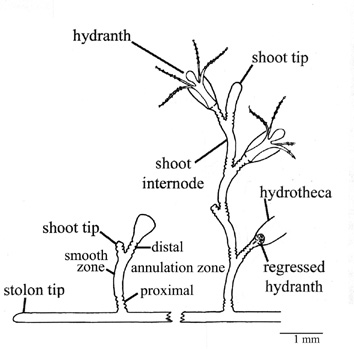 Schematic drawing of a colony
of Laomedea flexuosa
Schematic drawing of a colony
of Laomedea flexuosa