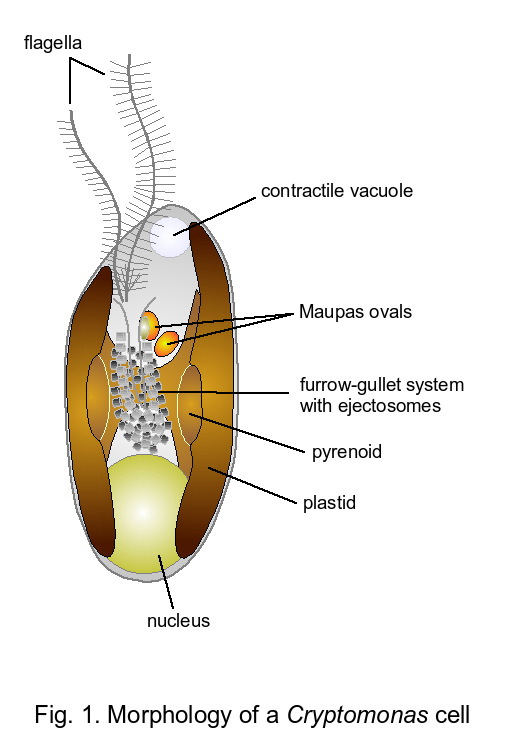
Cryptophyte Morphology
Cryptophytes are unicellular algae with cell lengths from around 3 (Hemiselmis species) up to 50μm (Cryptomonas curvata). The cells contain one to two plastids per cell and propel themselves through the water with two unequal flagella. One flagellum, usually the longer one, has two opposite rows of flagellar hairs (Hibberd et al. 1971). These stiff flagellar hairs cause a reversal of thrust, i.e. with the flagellum pointing in swimming direction, the cell is pulled through the water (= locomotoric flagellum; Sleigh 1991). The shorter flagellum shows only one row of flagellar hairs, which are attached in a different way than the locomotoric flagellum (Hibberd et al. 1971). In addition, delicate rosulate scales cover both flagella. Cryptophyte cells are more or less stereospecifically asymmetric. The flagella insert subapically in the vestibule of an invagination often shifted to the cell's right, whereas the cell apex is shifted to the left (Fig. 1).
Several Features are Unique to the Cryptophytes:
- A cell invagination, either a tubular gullet, a furrow, a combination of furrow and gullet or a groove, lined with explosive structures termed ejecto- or ejectisomes (Morrall and Greenwood 1980).
- The ejectosomes: two connected ribbons of different sizes, that are rolled up and under tension. They become discharged upon mechanical or chemical stress irritating the cells (Morrall and Greenwood 1980).
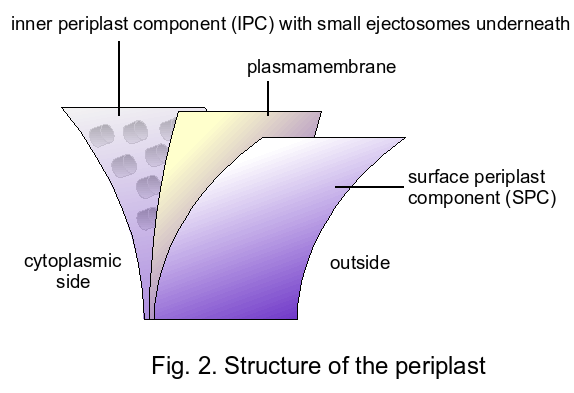
- The periplast: a sandwich-layered structure surrounding the cells that consists of proteinaceous inner and surface periplast components with the plasmamembrane in between (Brett et al. 1994; Fig. 2).
- A so-called complex plastid surrounded by four instead of only two membranes (Gillott and Gibbs 1980).
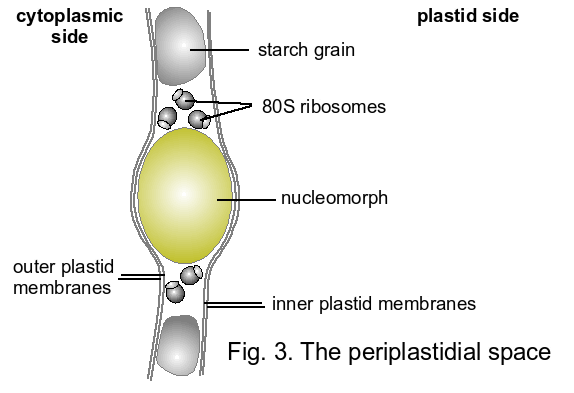
- The periplastidial space between the two inner and the two outer plastid membranes contains starch grains, 80S ribosomes and a second, but highly reduced nucleus termed the nucleomorph (Greenwood 1974; Gillott and Gibbs 1980; McFadden et al. 1994b; Douglas et al. 2001; Fig. 3).
- A light harvesting complex consisting of one type of either red or blue phycobiliprotein not attached to the outside of the thylakoids, but residing densely packed in the thylakoid lumina (Gantt et al. 1971).
The aplastidic cryptophyte genus Goniomonas lacks all features associated with the complex plastid of the photosynthetic cryptophytes: a periplastidial compartment, starch synthesis and a nucleomorph (Mignot 1965). It differs from plastid-containing cryptophytes also in cell shape (flattened in lateral plane), in cell invagination(s) and in flagellar structure. In addition to an invagination with a line of ejectosomes probably corresponding to the invagination found in photosynthetic cryptophytes, a cytostome is present (Mignot 1965). Also its flagella are different. One flagellum shows one row of stiff but bent flagellar hairs, whereas no such hairs are found on the other flagellum (Kugrens et al. 1987). Both flagella are covered by many thin and long fibrillar hairs not found in other cryptophytes.
Because of the mixture of photosynthetic and heterotrophic taxa, the cryptophytes are also refered to as cryptomonads. They can be classified either according to the International Code of Nomenclature for algae, fungi and plants (ICN) or according to the International Code of Zoological Nomenclature (ICZN).
Ultrastructural characters such as differences in periplast structures, in types of cell invagination, in flagellar apparatus ultrastructure and/or in the position of the nucleomorph have been used to define cryptophyte genera (Table 2). In addition, seven different types of biliproteins have been found; each clonal culture produces only one type of biliprotein. One new genus and species, Proteomonas sulcata, was found to be dimorphic in a clonal culture (Hill and Wetherbee 1986). Some cells had hexagonal inner periplast plates and non-keeled rhizostyles, whereas others had sheet-like inner periplast components and keeled rhizostyles (microtubular flagellar roots directed posteriorly). Since Hill and Wetherbee detected different amounts of DNA in the cell nuclei, they termed the different cell types haplomorph and diplomorph (Table 2) and assumed the presence of a dimorphic life history with sexual reproduction in Proteomonas sulcata. Similar to Proteomonas sulcata, evidence for dimorphism has been found in the freshwater genus Cryptomonas (Hoef-Emden and Melkonian 2003).