Evolutionary History: Four Genomes in a Cell
Cryptophytes resemble Russian dolls. Due to a successive series of endosymbiotic events, at least four different formerly separate organisms became nested within each other to combine to one lifeform (McFadden 2001; Reyes-Prieto et al. 2007; Fig. 1). Each of the organisms contributed its genome, thus four different genomes are present in a cryptophyte cell.
- The Presumed Endosymbiotic Events in Chronological Order
- Evolution of the Unique Cryptophyte Biliproteins
- Closest Relatives
The Presumed Endosymbiotic Events in Chronological Order
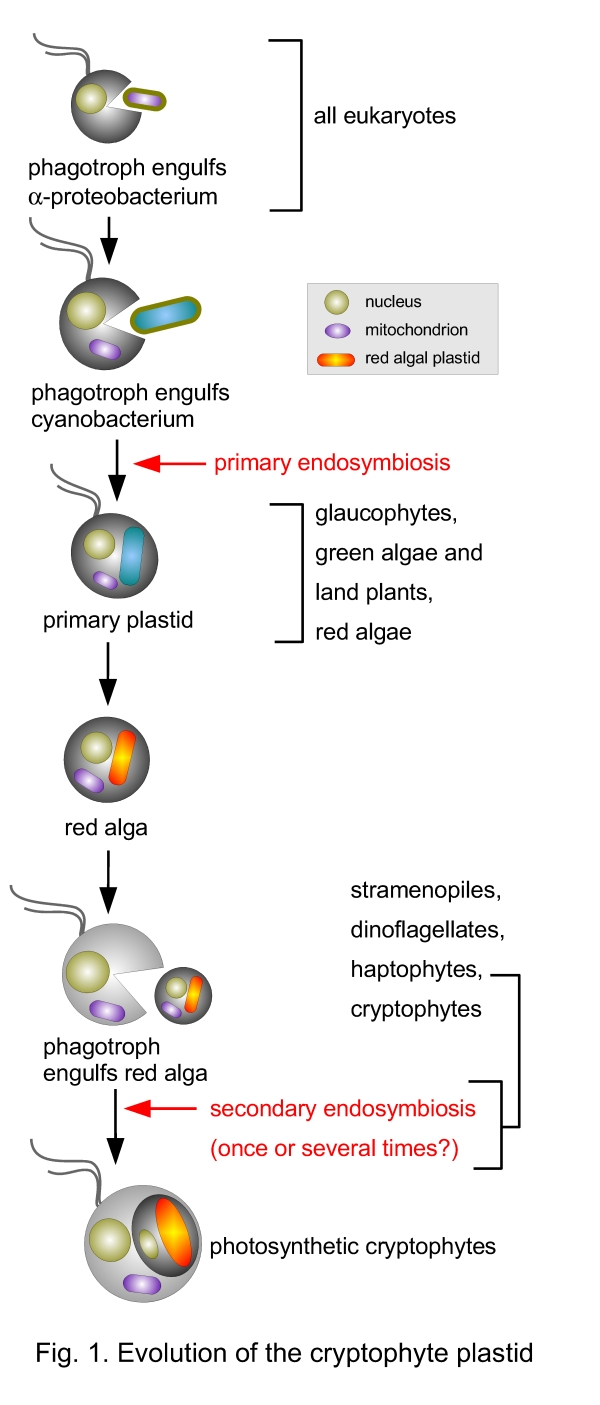
- A phagotrophic eukaryotic cell engulfed an α-proteobacterium. Genes from the circular bacterial genome were either lost or have been transferred to the host cell nucleus and the formerly autonomous bacterium became a cell organelle, the mitochondrion. This type of endosymbiosis is common to all extant eukaryotic cells and enabled them to respirate. In amitochondriate protists mitochondria have been lost secondarily (Homer et al. 1996; Germot et al. 1996).
- A phagotrophic eukaryotic cell containing already a mitochondrion (or several mitochondria) engulfed a cyanobacterium, a bacterium that can do oxygenic photosynthesis (fixing carbon dioxide during photosynthesis and releasing oxygene). The cyanobacterium also became a cell organelle, the plastid. It has a circular, but highly reduced genome and is surrounded by two membranes. This type of endosymbiosis happened in the common ancestor to all plants. The Plantae comprise the green algae, the land plants, the red algae and the glaucophytes (Reyes-Prieto et al. 2007).
- A phagotrophic cryptophyte (the host cell) engulfed a red alga and, similar to the endosymbiotic events listed above, instead of digesting it, the red alga became a cell organelle, a complex plastid surrounded by four membranes. The mitochondria of the red alga have been lost, whereas the host cell has kept its mitochondrion. The nucleus of the red alga has been extremely reduced and became the nucleomorph. Similar to the primary endosymbioses leading to mitochondria and primary plastids, a massive gene transfer to the host cell nucleus took place (Douglas and Penny 1999; Douglas et al. 2001; Deane et al. 2000).
Thus, a photosynthetic cryptophyte cell contains two linear eukaryotic genomes (host cell nucleus with the majority of DNA and nucleomorph = former red algal nucleus) and two circular prokaryotic genomes (a former α-proteobacterial and a former cyanobacterial genome). Red algae produce starch as an energy storage in the cytoplasm, not in the plastid as in green algae or land plants. Thus, it can be deduced from the eukaryotic ribosomes and the starch grains between the two inner and the two outer plastid membranes, that the periplastidial space corresponds to the former cytoplasm of the engulfed red alga. Complete nucleomorph genomes have been sequenced from Guillardia theta and Hemiselmis andersenii (Douglas et al. 2001; Lane et al. 2007) and two complete plastid genomes are available (Guillardia theta and Rhodomonas salina; Douglas and Penny 1999; Khan et al. 2007). Also a complete mitochondrial genome of a Rhodomonas salina strain has been sequenced (Hauth et al. 2005).
Cryptophytes are not the only algae with complex plastids derived from a secondary endosymbiosis. Complex plastids with more than two surrounding membranes are also found in stramenopile algae (brown algae, diatoms, eustigmatophytes, golden algae, raphidophytes), in photosynthetic dinoflagellates, in haptophytes, in euglenoids or in chlorarachniophytes (Reyes-Prieto et al. 2007). Only in the chlorarachniophytes, amoebae with a complex plastid of green algal origin, also a nucleomorph is found – in this case the former nucleus of a green alga (Gilson et al. 2006).
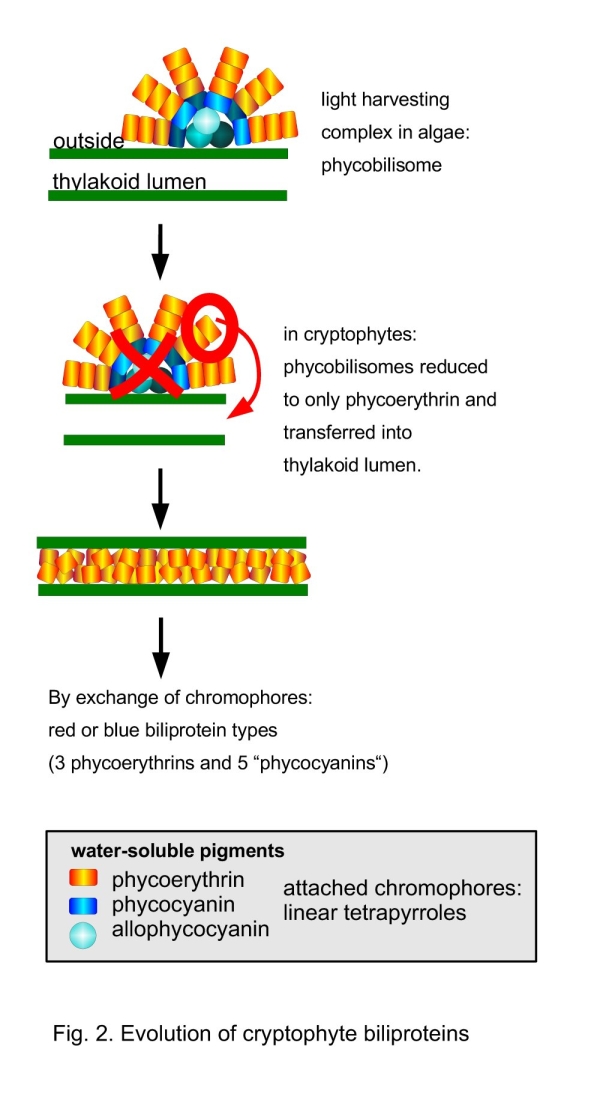
Evolution of the Unique Cryptophyte Biliproteins
The plastids of red algae and glaucophytes inherited their light harvesting complexes – the phycobilisomes – from the former cyanobacterial endosymbiont (MacColl 1998; Stirewalt et al. 1995; Ohta et al. 2003). Like the chlorophyll-containing light harvesting complexes they are attached to the outside of the plastid endomembrane system (the thylakoids). Phycobilisomes comprise several biliproteins (allophycocyanins, phycocyanins and in many red algae also phycoerythrins). Allophycocyanins and phycocyanins are blueish, phycoerythrins red. In cryptophytes, all genes for allophycocyanins and phycocyanins seem to have been lost and the remaining phycoerythrin has been transferred into the thylakoid lumina (Gantt et al. 1971; Apt et al. 1995; Fig. 2). Its subunit structure changed from (αβ)3 or (αβ)6 to (αβ)(α'β) (Glazer and Wedemayer 1995). The gene for the β-subunit is plastid-encoded, whereas the genes for the α-subunits, comprising at least two different genes, have been transferred to the nucleus (Gould et al. 2007). By exchange of the tetrapyrrole chromophores, the phycoerythrin changed colour and absorption spectrum in several cryptophyte lineages. As a result, not only different red, but also blue types of phycoerythrins (cryptophyte “phycocyanins”) evolved (Hill and Rowan 1989; Glazer and Wedemayer 1995). Despite of the profound changes in structure and of the the peculiar intra-thylakoidal organisation, the cryptophyte biliproteins seem to operate as active light-harvesting complexes (Doust et al. 2006).
Closest Relatives
The next related group to the cryptophytes apparently are the katablepharids, phagotrophic flagellates with explosive organelles resembling modified cryptophyte ejectosomes (Okamoto and Inouye 2005). Since the haptophytes grouped in previous phylogenetic analyses as a sister to cryptophytes and katablepharids, the three taxa have been united under the name Hacrobia (Patron et al. 2007; Okamoto et al. 2009). The chromalveolate hypothesis assumes that Hacrobia, Stramenopiles, ciliates, dinoflagellates and Apicomplexa are monophyletic and can be merged in one supergroup or kingdom Chromalveolata (Cavalier-Smith 1999). Whereas in phylogenetic trees inferred from supermatrices of plastid-encoded or nuclear-encoded plastid-targeted genes, support for the chromalveolate hypothesis is found, phylogenies inferred from genes of the host cell contradict this hypothesis (Yoon et al. 2002; Harper and Keeling 2003; Bachvaroff et al. 2005; Hackett et al. 2007; Yoon et al. 2008).