What is a Species?
By Taxonomy
Taxonomy is the science of the classification of lifeforms. The rules that have to be followed for a valid description of taxa are laid down in nomenclature codes by the respective taxonomic societies. Taxonomy is governed by long-term traditions, therefore the assignment of organisms to a nomenclature code does not always make sense from an evolutionary point of view (Table 1). Irrespective of their evolutionary history, heterotrophic and photoautotrophic eukaryotes are still treated by tradition in general as "animals" or "plants", respectively, or under both codes with sometimes contradictive outcome. Although for prokaryotes a special bacterial nomenclature code has been established, the Cyanobacteria – the group of bacteria that invented oxygenic photosynthesis – are often still treated as "plants" under the botanical nomenclature code. Also the rules for the classification Fungi, the closest relatives of animals and choanoflagellates, by tradition have been part of the botanical code.
Table 1. Current Classification of Eukaryotes and their Assignment to Nomenclature Codes
| Kingdom or Supergroup | Group of Organisms (Division, Subdivision, Phylum or Class) | Assigned Nomenclature Code |
| Opisthokonta | Metazoa (animals), Choanoflagellida | ICZN |
| Fungi | ICN | |
| Plantae/Archaeplastida | Glaucoplantae, Rhodoplantae (red algae), Viridiplantae (green algae and embryophytes) | ICN |
| Chromalveolata | Stramenopiles (heterokonts): Bacillariophyta (diatoms), Dictyochophyceae (silicoflagellates), Eustigmatophyceae, Oomycetes, Raphidophyceae, Phaeophyceae (brown algae) | ICN |
| Stramenopiles: Chrysophyta (golden algae) | ICN and ICZN | |
| Stramenopiles: Bicosoecida, Labyrinthulida | ICZN | |
| Alveolata: Dinophyceae (dinoflagellates) | ICN and ICZN | |
| Alveolata: Apicomplexa (parasites causing malaria, toxoplasmosis etc.) | ICZN | |
| Alveolata: Ciliophora (ciliates) | ICZN | |
| Hacrobia | Haptophyta (prymnesiophytes) | ICN |
| Cryptophyta | ICN and ICZN | |
| Katablepharidae (katablepharids) | ICZN and ICN | |
| Rhizaria | all to the exclusion of Paulinella chromatophora and Chlorarachniophyceae | ICZN |
| Cercozoa: Paulinella chromatophora | ICN and ICZN | |
| Cercozoa: Chlorarachniophyceae | ICN | |
| Amoebozoa | all groups | ICZN |
| Excavata | all groups to the exclusion of Euglenida | ICZN |
| Euglenida | ICZN and ICN |
Note: Table 1 does not represent an exhaustive listing of all sub-groups within the eukaryotic kingdoms. Abbreviations: ICN, International Code of Nomenclature for algae, fungi and plants (Melbourne Code); ICZN, International Code of Zoological Nomenclature; non-eukaryotic codes of nomenclature not mentioned in Table 1: International Code of Nomenclature of Bacteria (ICNB), The International Code of Virus Classification and Nomenclature (ICNV)
In the nomenclature codes the ranks of classification and the correct endings of rank names are defined. Most of the major ranks are common to all classification systems. The species is always the lowest major rank, i.e. the smallest classification unit (regnum = kingdom; divisio/phylum = division/stem; classis = class; ordo = order; familia = family; genus; species). Minor ranks below are subspecies and variety. A species name follows the binomial nomenclature established by the Swedish botanist Carl Linnaeus (also known as Carl von Linné) for plants in 1753 and for animals in 1758. A species name always consists of a combination of a genus name with an epithet. To be valid, the species description has to follow the rules of the respective nomenclature code. The botanical code e.g. requests the mentioning of the locality and a specimen (the holotype) has to be submitted as a voucher material to a public collection or museum to allow access for future research. Many people think that classification and naming and/or the description of species is well-established and sound science that can be relied upon. Classification systems, however, are volatile. They have been subject to many changes during the course of time and the definition of species often is anything but clear. Neither the botanical nor the zoological code tell whether it makes sense to describe an organism as a new species by using this or that character, nor do they mention DNA as a means of identification.
Different Species Concepts
 The morphological or morphospecies concept is the oldest species
concept and uses differences in morphological characters to distinguish species. In former times species
were considered as inalterable units.
After Charles Darwin published his book "On the Origin of Species by Means of Natural Selection" (1859),
the morphospecies concept has been applied under the assumption that similar to identical morphological
characters would reflect relatedness and therefore result in a natural systematics.
The morphological or morphospecies concept is the oldest species
concept and uses differences in morphological characters to distinguish species. In former times species
were considered as inalterable units.
After Charles Darwin published his book "On the Origin of Species by Means of Natural Selection" (1859),
the morphospecies concept has been applied under the assumption that similar to identical morphological
characters would reflect relatedness and therefore result in a natural systematics.
Ernst Mayr invented the biological species concept in 1942. A biological species is a group of individuals that can interbreed and produce fertile offspring to the exclusion of others. To determine boundaries of biological species, time-consuming mating experiments are necessary. Provided mating experiments are possible, a species defined by the biological species concept is reproducible and represents the smallest taxonomic and evolutionary unit. Biological species are therefore considered the optimal species definition. However, especially in plants, but also in some animals the biological species concept may not work out due to incomplete separation of evolutionary lineages.
The phylogenetic species concept uses molecular markers – usually DNA sequences – to identify closely related taxa. It is e.g. the standard species concept in bacteria, but has been tested also for eukaryotic taxa (Leliaert et al. 2009). This species concept lacks support by botanical and zoological nomenclature codes, because it represents a DNA-based taxonomy. Some time ago the Phylocode has been introduced as an alternative to the morphology-based botanical and zoological nomenclature codes, but has not been widely accepted yet. The Phylocode uses monophyletic clades as taxonomic units, but does not implement the traditional classification ranks.
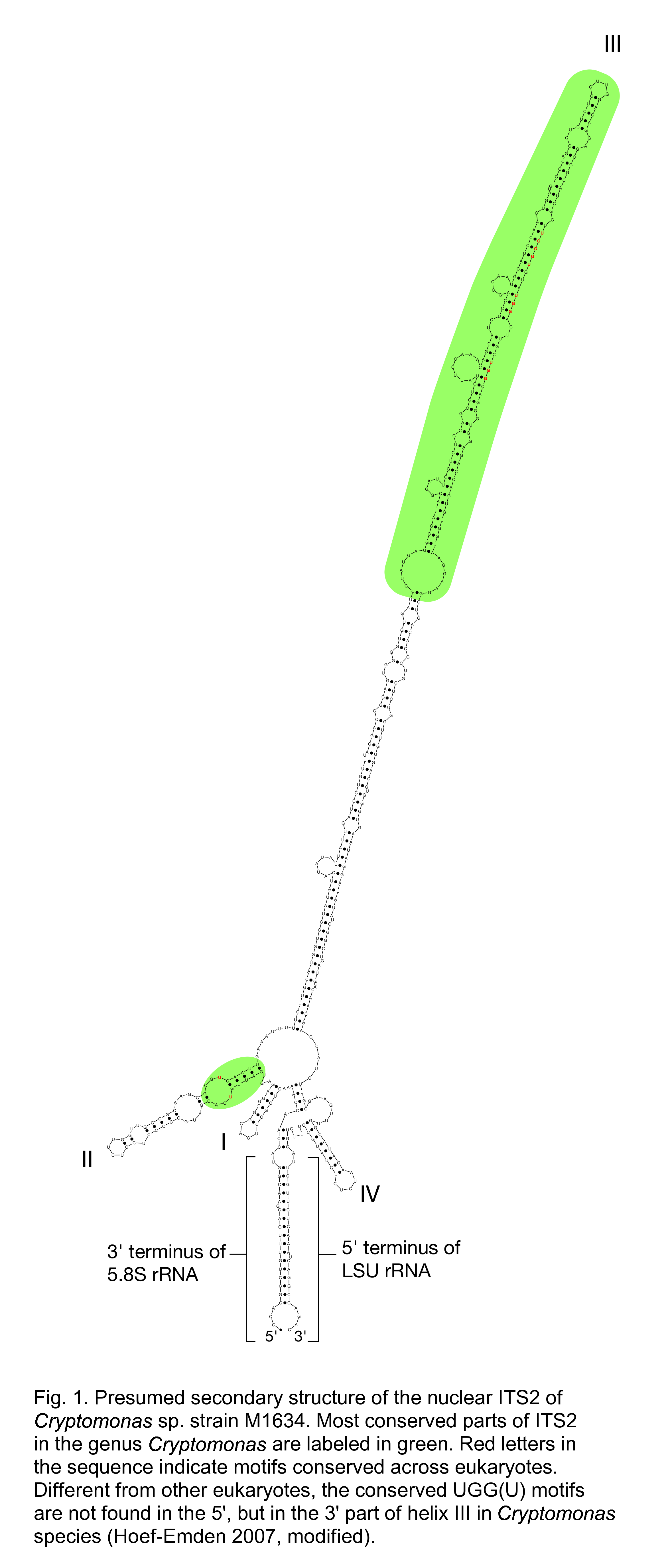
CBC is the abbreviation for Compensatory Base pair Changes. The CBC clade concept is related to the phylogenetic species concept, but can only be applied to eukaryotic organisms, because it is based on the secondary structure of the highly variable internal transcribed spacer 2 (ITS2) of the eukaryotic ribosomal operon. The ITS2 separates 5.8S and 28S (or LSU) rRNA genes and has to fold up prior to being excised from the huge primary transcript. In a correctly folded ITS2, 3' and 5' termini of 5.8S and LSU rRNA, respectively, pair to form a stem from which the ITS2 loops out – in most cases – into four helices (Mai and Coleman 1997; Schultz et al. 2005; Fig. 1). Helices II and III contain important parts for recognition and splicing (e.g. the initial C2 cleavage site close to the tip of helix III) (Côté and Peculis 2001; Schultz et al. 2005). Although ITS2 sequences are so highly variable, that they usually cannot be aligned even across related genera, several sequence elements have been found in helices II and III, that are conserved across eukaryotes (Schultz et al. 2005).
In a comparison of mating experiments with secondary structure analysis of ITS2 in green algae of the order Volvocales, one complete exchange of a base pair in the proximal part of helix II or across helix III proved to be correlated with sexual incompatiblity of the gametes of the tested algal strains, whereas helices I and IV could be quite variable (Coleman 2000). Since then, several studies confirmed this correlation between ITS2 secondary structure and mating failure also for other eukaryotes (summarised in Coleman 2009). In the latter publication the region that has to be identical across putative biological species has been even reduced to the 30 most conserved nucleotides at the 5' terminus of helix III. Annette Coleman considered as a possible explanation for this phenomenon, that high evolutionary rates in mating-related genes result very soon in gamete incompatibility, whereas CBCs in helix III need more time to happen (Coleman 2009). Thus, compensatory base pair changes in the helix III are safe predictors for sexual incompatibility between evolutionary lineages, but the reverse direction, assuming that the absence of CBCs indicates sexual compatibility may not necessarily be true.
All species concepts have their shortcomings and not all species concepts can be used for all protists.